INTRODUCTION
In the past, lactate was considered merely a by-product, with early studies reporting that it caused a reduction in pH and decreased exercise performance [1-3]. However, recent studies on lactate have revealed its role in protecting against metabolic acidosis, and have reconsidered it as a potential fuel source for exercising muscle [4-6]. Representatively, the inter- and intracellular lactate shuttle theory [such as monocarboxylate transporters (MCTs)] present lactate as a recycling energy resource for the body during or after exercise, as well as during resting conditions [7-9]. The change in perception suggests that exogenous lactate can be used as a supplement to exercise.
A recently published study reported that combined intake of exogenous lactate and caffeine under resting conditions induced fat oxidation and increased hepatic glycogen concentration in rats [10]. Notably, the effects were similar to those in the lactate-only intake group in this study. Previously, we confirmed metabolic-related variables and reported that 2 g/kg exogenous lactate intake increased the factors involved in fat oxidation in the soleus muscles of rats [11]. Moreover, Kitaoka et al. [12] also reported that an intraperitoneal injection of 1 g/kg lactate enhanced the transcription of peroxisome proliferator-activated receptor-gamma coactivator 1 (PGC1)-alpha, which is related to mitochondrial biogenesis, after 3 h; Cerda‐Kohler et al. [13] demonstrated that 1 g/kg lactate injection phosphorylated the controlling skeletal muscle fat oxidation factor of adenosine monophosphate-activated protein kinase (AMPK). The studies revealed that lactate treatment in the resting state enhanced fat oxidation and glycogen synthesis, and has the potential to upregulate energy utilization efficiency during exercise.
Furthermore, Takahashi et al. [14] reported that 4 weeks of oral intake of 5 g/kg lactate induced mitochondrial enzyme activity by increasing citrate synthase and cytochrome c oxidase activation in the oxidative muscles of mice and synergized with endurance exercise (20 m/min for 30 min). Hoshino et al. [15] confirmed that at 3 weeks post-exercise (6 days/week of 40 min treadmill running at 25 m/min), lactate intake increased glycogen concentration and enhanced MCT1 protein levels, which led to an influx of lactate into the oxidative tissue of the tibialis anterior muscle of mice. In addition, the administration of a 1 g/kg lactate-based compound combined with low-intensity exercise activated anabolic signals and increased gastrocnemius and weights of tibialis muscle in rats [16]. When the aforementioned information is considered as a whole, exogenous lactate can serve as an exercise supplement, leading to the hypothesis that lactate intake after endurance exercise can enhance exercise performance through exercise-induced adaptation. However, previous studies have not confirmed the effects of lactate intake on exercise performance by evaluating metabolism during exercise.
Therefore, to determine whether repetitive exogenous lactate intake with or without exercise increased exercise performance, the study investigated the combined effects of 4 weeks of exogenous lactate intake and endurance exercise on metabolism in VO2max measurement.
METHODS
Animal care
Six-week-old male ICR mice (n =28) were purchased from Orient Bio Inc. (Seongnam, South Korea). All mice were housed in standard plastic cages under controlled conditions of humidity (50%) and temperature (23 ± 1 °C) with an alternating 12 h light-dark cycle. The mice were acclimated to the laboratory housing conditions for 7 days. The study was approved by the Institutional Animal Care and Use Committee of Konkuk University (no. KU19149).
Protocol
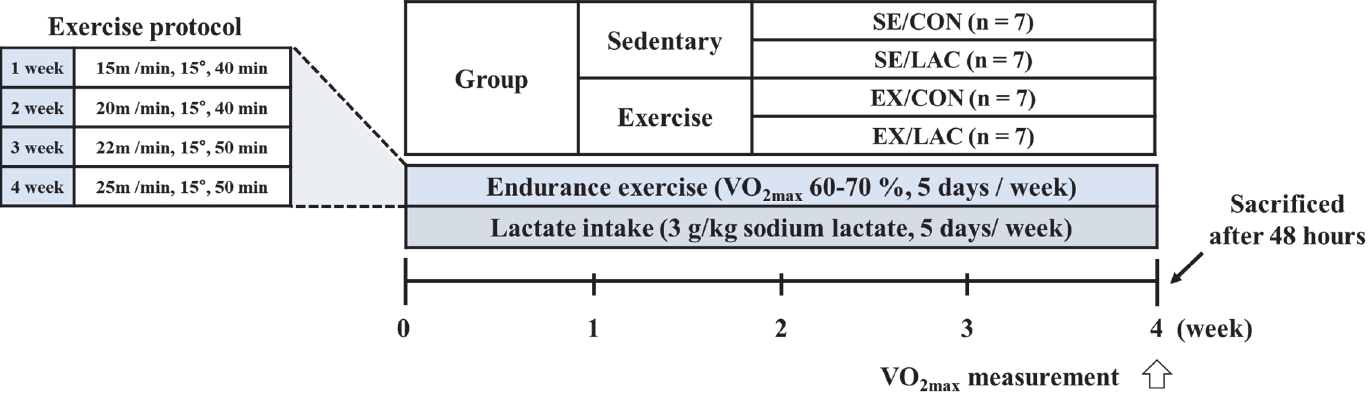
The mice were randomly divided into two groups (sedentary and exercise) and further subdivided into the following four groups: control (SE/CON, vehicle intake), lactate intake (SE/LAC), exercise (EX/CON, endurance exercise with vehicle intake), and lactate intake with endurance exercise (EX/LAC). The exercise groups (EX/CON and EX/LAC) underwent treadmill training (Daejong Systems, Korea) at a fixed intensity (15 m/min, 8 ° slope) for 2 days. Thereafter, endurance exercise was performed 5 days a week, and exercise intensity was gradually increased to achieve a 60-70% maximal oxygen uptake (VO2max). The protocols were modified from those of Hwang et al. [17] as follows: 1 week, 15 m/min, 8 ° slope, 40 min; 2 weeks, 20 m/min, 8 ° slope, 40 min; 3 weeks, 22 m/min, 8 ° slope, 50 min; and 4 weeks, 25 m/min, 8 ° slope, 50 min. The lactate intake groups (SE/LAC and EX/LAC) were orally administered sodium lactate (3 g/kg) immediately after exercise [18,19], meanwhile, the vehicle intake groups (SE/CON and EX/CON) were treated with the same volume of saline. After 48 h of VO2max measurement, the mice were sacrificed using intraperitoneal anesthesia with 10 μL/g of 1.25% avertin, and three abdominal fat samples (epididymal, perirenal, and mesenteric) were collected. The detailed study design is illustrated in Figure 1.
VO2max measurement
All the mice were subjected to VO2max measurements in a metabolic chamber during exercise after the experimental period using an open-circuit calorimetry system. VO2max measurements were performed by the study protocol. Briefly, the initial slope and speed were set to 0 ° and 14 m/min, respectively, and then increased by 2 ° and 2 m/min, respectively, every 2 min.
Gas analysis
Respiratory gas analysis was performed using open-circuit equipment in accordance with the previous studies [10,18]. Oxygen uptake and carbon dioxide (CO2) production were measured using a mass analyzer (model RL-600; Alco System, Chiba, Japan) and a switching system (model ANI6-A-S; Alco System Chiba, Japan). The flow rate was maintained at 3 L/min. Oxygen uptake and CO2 production were used to calculate the respiratory exchange rate (RER), carbohydrate oxidation, and fat oxidation.
Statistical analysis
We used the IBM SPSS Statistics 25 software to analyze all data. Significant differences in body weight, abdominal fat mass, VO2max and time to exhaustion (TTE) were determined using a two-way analysis of variance followed by the least significant difference test. Significant differences in RER, fat oxidation, and carbohydrate oxidation were determined using two-way repeated measures analysis of variance. Values of p < 0.05 were considered statistically significant. Body weight, abdominal fat mass, VO2max and TTE results were presented as mean ± standard deviation (n = 7) and RER, fat oxidation, and carbohydrate oxidation were presented as mean ± standard error of the mean (n = 7).
RESULTS
Body weight and abdominal fat mass
After 4 weeks, body weight and abdominal fat mass were measured, as presented in Table 1. Final body weights were not significantly different between the groups for the experimental period. The abdominal fat mass was measured after euthanasia. Lactate intake tended to decrease abdominal fat mass; however, there were no significant differences among the groups.
Maximal oxygen consumption (VO2max) and TTE
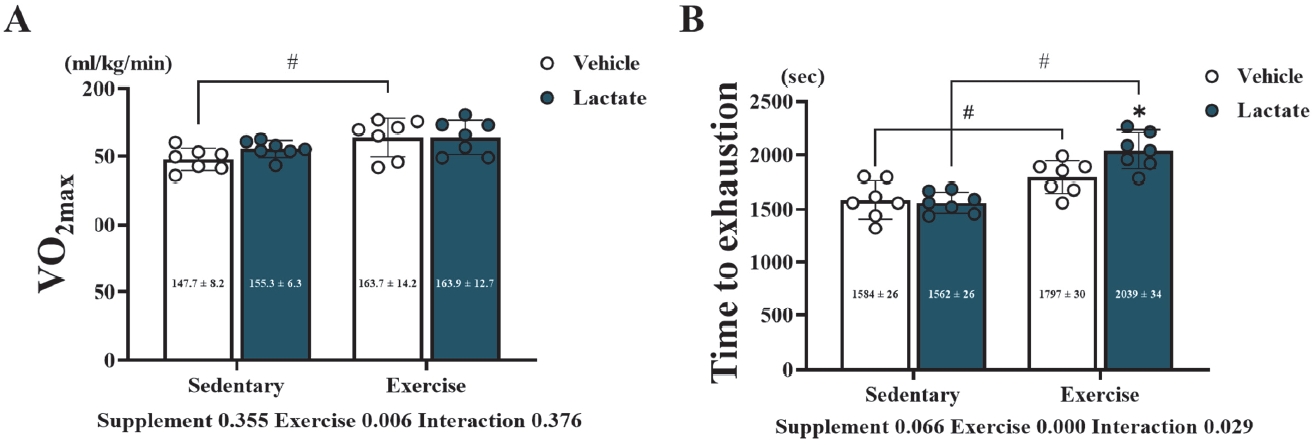
To confirm the effects of 4 weeks of lactate intake during endurance exercise on exercise performance, VO2max measurements were performed [20]. The VO2max was measured for the peak value of each mouse and later averaged. The VO2max exhibited only an exercise-dependent effect (p = 0.006); and in the vehicle treatment group, VO2max of the SE/CON group was higher than that of the EX/CON group (p < 0.05) (Figure 2A). The TTE was also evaluated and averaged. The TTE also displayed an exercise-dependent effect (p = 0.000) along with an interaction effect (p = 0.029). After the post hoc test, the TTE values in the exercise groups (EX/CON and EX/LAC) were higher than those in the sedentary groups (SE/CON and SE/LAC) (p < 0.05); and the TTE in the EX/LAC was higher than that in the SE/LAC (p < 0.05).
Change in energy substrate utilization during VO2max measurement
A significant time effect (p = 0.000), and interaction (p = 0.024) were observed in RER (Figure 3A). The SE/CON group generally displayed the highest RERs in VO2max measurement. Although the SE/LAC group had lower RERs than the SE/CON group, the TTE values were similar. The EX/CON and EX/LAC groups demonstrated a similar increase in RER, but the increase in the EX/LAC group was relatively higher. Similarly, based on the fat oxidation results (time effect, p = 0.000; interaction, p = 0.026), the SE/CON group generally exhibited the lowest fat oxidation levels (Figure 3B). The results of carbohydrate oxidation (Figure 3C) displayed a significant difference only in the time effect (p = 0.000). All groups exhibited similar increases in VO2max measurements.
DISCUSSION
The purpose of the present study was to determine whether 4 weeks of exogenous lactate intake, with or without endurance exercise, increased exercise performance. The results confirmed that 4 weeks of lactate intake improved fat oxidation and increased VO2max. Moreover, the results also established that 4 weeks lactate intake with endurance exercise enhanced exercise performance.
The results of this study revealed that the body weight did not differ among the four groups. Furthermore, abdominal fat mass tended to decrease after 4 weeks of lactate intake, but it was not significantly different among the groups. One reason for these results may be the different rodent models used in the present study. Normal and obese mice have different energy metabolisms, and obese mice have higher fat metabolism than normal mice [21,22]. Previous studies in which lactate treatment decreased fat mass used obese mice and suggested that lactate treatment affected obesity more than in normal mice by enhancing fat oxidation. Another possibility is that the study period of 4 weeks was insufficient to decrease fat mass. Moreover, previous studies also reported that 12 weeks of lactate administration alleviated the accumulation of fat mass [23] and that 5 weeks of lactate-based compound treatment and voluntary exercise decreased fat mass [16]. However, at 3 weeks post-exercise, lactate treatment tended to result in lower epididymal fat mass, and 4 weeks of lactate treatment did not affect the body weight [14,15]. Therefore, the experimental study period was shorter than that in previous studies, suggesting that 4 weeks of lactate treatment is insufficient to decrease fat mass.
The study aimed to determine whether lactate intake with or without endurance exercise improved exercise performance. As mentioned in previous studies, exogenous lactate intake has the potential to increase fat oxidation under resting conditions [10,11,12,23], which suggests that lactate intake may enhance exercise performance. Furthermore, this study used VO2max measurements to evaluate exercise performance. The results demonstrated that the SE/LAC group had higher fat oxidation than the SE/CON group in VO2max measurement, and it was speculated that 4 weeks of lactate intake could upregulate fat oxidation during exercise. In addition, previous studies have confirmed that lactate stimulates AMPK [13,24] and PGC1-alpha signaling [12,25], which are major factors in mitochondrial biogenesis, support the results exhibiting increased fat oxidation during exercise. However, the maximum exercise time did not differ between the SE/CON and SE/LAC groups. This suggests that, although the ability to utilize fat increased with lactate intake, muscle function did not develop as exercise was not performed. In addition, regarding VO2max, the VO2max measurements of the exercise groups (EX/CON and EX/LAC) were higher than those of the sedentary groups (SE/CON and SE/LAC). Notably, the study confirmed that lactate intake and endurance exercise had no additional effects on fat oxidation during exercise but enhanced TTE (EX/CON < EX/LAC) in VO2max measurement. Takahashi et al. [14] investigated the effect of 4 weeks of lactate intake before endurance exercise and confirmed that it enhanced the activation of mitochondrial enzymes, whereas Hoshino et al. [15] reported that at 3 weeks post-exercise, lactate intake increased the protein expression of MCT1 and upregulated glycogen recovery and storage in skeletal muscle. Interestingly, the two studies established that lactate treatment along with exercise had no additional effect on the fat-oxidation factors (AMPK, acetyl-coenzyme A carboxylase, and cluster of differentiation 36). Additionally, the studies suggest that lactate intake during endurance exercise may enhance the exercise capacity by rapidly supplying energy rather than by increasing fat oxidation, or by using lactate as an energy resource during exercise. Furthermore, several previous studies have reported that lactate-based mixed compounds and exercise increase skeletal muscle mass in both normal [26] and obese rats [16]. This suggests that lactate intake combined with endurance exercise induces additional effects on skeletal muscle hypertrophy, suggesting that it may positively affect muscle function. However, a limitation of the present study is that it lacked a reference on the lactate dose, treatment methods, and timing. The study used an oral intake of 3 g/kg lactate after endurance exercise. However, previous studies on lactate treatment have used a wide range of doses (1-5 g/kg) and various treatment methods (injection, oral intake, etc.). Moreover, the appropriate timing of lactate treatment with exercise was not established (pre-, post-, or separately). Nevertheless, the study confirmed that lactate intake, when consumed as an exercise supplement, improved exercise performance, which is speculated to be due to the multiple effects exerted by lactate (fat oxidation, mitochondrial biogenesis, muscle hypertrophy, etc.).
In conclusion, although a decrease in fat mass was not observed with lactate intake, the present study suggests that exogenous lactate has the potential to improve exercise performance as an exercise supplement. However, the present study has a limitation in that fragmentary results were confirmed. Therefore, to use lactate as an exercise supplement, further studies are needed to reveal the details of the mechanisms of lactate intake and exercise and to include the study of lactate levels, treatment methods, and timing.