



INTRODUCTION
The blood vessels of humans are typically in a static state. Except for the early developmental period, scarring, and during the menstrual cycle, almost no angiogenesis occurs1. However, many recent studies have indicated that various conditions affect angiogenesis of blood vessels.
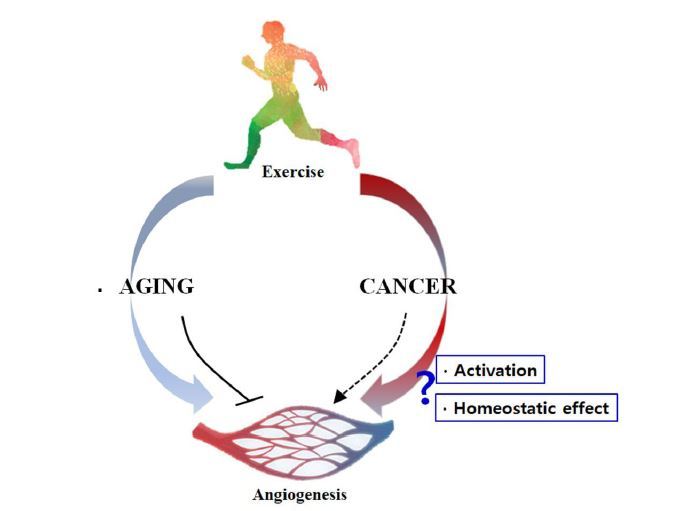
Especially, exercise and cancer affect angiogenesis, generally by promoting angiogenesis, in contrast to aging, which decreases angiogenesis2-4. Angiogenesis can be viewed as a phenomenon linking exercise, aging, and cancer, and the same time being affected by these aspects1-6. However, although exercise, aging, and cancer can be concurrent, little research has been done on them.
Factors that regulate angiogenesis are termed angiogenic factors; they are important in inducing the formation of blood vessels in the quiescent period, and involves cytokines that include vascular endothelial growth factor (VEGF), fibroblast growth factor (FGF), and hypoxia-inducible factors (HIFs)7,8. Endothelial cells are directly involved in angiogenesis and act directly with angiogenic factors via tyrosine kinase receptors to regulate angiogenesis9. In addition, skeletal muscle cells secrete an angiogenic factor that affects angiogenesis10. In particular, the vascular molecules that angiogenic factors directly interact with provide nutrient support to skeletal muscles11. Thus, angiogenic factors are very important in skeletal muscle.
This review provides and explanation of the correlation between angiogenic factors, exercise, and aging in endothelial and skeletal muscle cells. In particular, we focus on studies that have examined the effect of exercise on endothelial cells, rather than skeletal muscle. The data may have significant meaning concerning angiogenesis in skeletal muscle. In addition, because cancer also affects angiogenesis, we examine the relationship between exercise and cancer in terms of angiogenesis.
Exercise and Angiogenesis
Exercise has a positive effect on long-term cardiovascular disease 12. In particular, exercise plays a major role in alleviating vascular resistance and stiffness13.
A number of in vitro, cell-based studies have been conducted to evaluate the changes in the pattern of angiogenesis pattern after exercise using endothelial cells to try to mimic the in vivo condition14,15. Endothelial cells are the major cells forming blood vessels16, and are influenced by angiogenic factors that bind to tyrosine kinase receptors9. The identification of angiogenesis-related molecules that mimic exercise in endothelial cells is directly related to endothelial vascular formation. An example is the influence of 5-aminoimidazole-4-carboxamide ribonucleotide (AICAR) on human umbilical vein endothelial cells (HUVEC) and rat myocardial microvascular endothelial cells (MMEC)14,15,17. In the case of HUVEC cells, AICAR treatment decreased the amount of fatty acids and lactate14, In case of MMEC, AICAR treatment increased AMPK and VEGF mRNA expression17. Bovine capillary endothelial (BCE) cells are also representative endothelial cells18. BCE cells were treated with soleus muscle extracts of rats treated with 15 weeks of exercise. Cell fluorescence was increased compared to the cells not treated with extracts. These results indicated that endothelial cell growth can be induced by exercise. The significance of this result is that exercise promotes angiogenesis by promoting the proliferation of endothelial cells that directly form blood vessels18.
C2C12 cells are representative skeletal muscle cells19. There are many ways to mimic the mechanism of exercise in C2C12 cells. The most commonly used methods include AICAR-based chemical methods. Other methods include physical stimulation of skeletal muscle cells through electrical stimulation20,21. As a result of treatment with AICAR, phosphorylation of AMPK and its subordinate signals were induced. Thus, the expression level of VEGF was confirmed by real-time PCR, The amount of VEGF expression was increased in proportion to the amount of AICAR. This result was almost identical to that obtained by insulin treatment. Treatment with AICAR pathway inhibitor and insulin pathway inhibitor confirmed that the mechanism of VEGF promoted by exercise and by insulin were different from those of AICAR and insulin22. VEGF expression using AICAR to mimic exercise resulted from the increased mRNA stability through AMPK, and VEGF expression by insulin was the result of increased transcription levels through phosphoinositol-3-kinase22. When electric pulse stimulation was applied to C2C12 cells, signals including extracellular signal-regulated kinase 1/2 and 5, c-Jun N-terminal kinase, and JNK and ATF, which are influenced by exercise, were activated as was glucose transporter type 423. When primary muscle cells were treated with EPS , the content of VEGF, which affects angiogenesis, was significantly increased. The increase was not immediate but occurred over the ensuing 24 hours. Fibroblast growth factor (FGF) increased only after EPS treatment compared to control group24. The observations suggest that the secretion of angiogenic factor in the muscles due to exercise continues both immediately after exercise and for some time after exercise (Table 1).
Table 1.
Exercise mimetic effects on cellular models.
| Cultured Cell Type | Exercise mimic | Molecular change |
|---|---|---|
|
HUVEC14 MMEC17 |
AICAR treatment |
Fatty acidsâ Lactateâ VEGF mRNAâ |
| BCE cells18 | Exercise treated mouseâs soleus muscle extract 18 | DNA fluorescenceâ |
| C2C12 cells | 0.5 mM AICAR22 | VEGF mRNA 1.5-foldâ |
| 2 mM AICAR22 | VEGF mRNA 2-foldâ | |
| EPS treatment23 |
Erk 5â Erk 1/2â JNKâ ATFâ GLUT4â |
|
| Primary muscle cell24 | EPS treatment |
VEGFâ FGFâ |
In an in vivo experiment, obese and normal weight male Zucker rats were exercised at increase exercise intensity five times a week for 8 weeks25. The level of VEGF-A was increased in the epididymal adipose tissues of all rats25. In this case, angiogenesis in the adipose tissue plays a role in inhibiting hypoxia and inflammation, mitigating cardiovascular disease26. In the case of 4-week-old Sprague-Dawley rats, mRNA expression levels of angiogenic factors that included VEGF, mammalian target of rapamycin (mTOR), and mTOR-C in the skeletal muscle increased after treadmill exercise27. To examine the effects of the resistance exercise, a clinical study was performed in which serum was collected and analyzed for 8 weeks after an exercise regimen. The angiogenic factors VEGF and angiopoietin 1 (Ang1) were significantly increased28. The in vivo and clinical data confirmed the inducing effect of angiogenesis by exercise and the appropriate exercise intensity for inducing angiogenesis (Table 2).
Table 2.
Exercise effects in in vivo models
| Model | Exercise | Molecular change |
|---|---|---|
| Male Zucker rat; Obesity, normal Model25 | 20 m/min for 60 min, 5 days each week for 8 weeks | In adipose tissue, VEGF-A levelâ |
| Sprague-Dawley rats27 | 5 days each week for 8 weeks, incremental treadmill exercise |
In skeletal muscle, VEGFâ mTOR-Câ mTORâ (mRNA expression) |
Aging and Angiogenesis
Aging is associated with diminished genetic processes, mitochondrial degeneration, cell death, and loss of enzyme activities 29. In particular, in some tissues associated with angiogenesis, VEGF, which is the most basic and dominant regulator, decreases with aging30. VEGF usually acts on endothelial nitric oxide synthase (eNOS) to induce vasodilation and angiogenesis. During senescence, eNOS activity decreases due to oxidative stress, decreased activity of sirtuin 1 (SIRT1), and decreased estrogen production6. In blood vessel, capillary density and formation, which are important in the removal of metabolic byproducts and oxygen transport, are directly affected by angiogenic factors31. We examined the angiogenesis patterns in human dermal microvascular endothelial cells (HMVECs) using aged and neonatal HMVECs, and found that capillary formation in aged HMVECs was reduced compared to neonatal cells, and that VEGF mRNA and protein levels were significantly decreased in aged HMVECs30.
Doxorubicin is used for chemotherapy in cancer patients. When administered to C2C12 cells, doxorubicin increases protein degradation through the production of reactive oxygen species (ROS) of mitochondria and induces catabolism32. Elevated ROS levels and increased protein degradation due to mitochondrial dysfunction in muscle may be a hallmark of aging33. Another study described that doxorubicin induces the production of urokinase receptor (uPAR), which regulates telomeric repeat binding factor 2 (TRF2), in turn causing ubiquitination and proteasomal degradation34. Therefore, by treating C2C12 cells with doxorubicin, it is possible to mimic the environment of aged C2C12 cells. Until now, there has been no comparative study of changes in VEGF, Ang1, platelet-derived growth factor (PDGF), and basic FGF (bFGF), but the overall effect of aging is expected to reduce angiogenic factors.
In vivo studies have shown that capillary density, which can be calculated as the capillary area per fiber area, decreases with age35. Comparison of the capillary density of 5, 13, and 25 month old rats with plantaris muscle has revealed the decreased capillary area with aging36. Another study compared hind limb capillary densities of 12-week-old mice and 24-month-old mice to investigate the influence of aging. The capillary density of the 24-month-old mice was twice that of the younger mice37.
In a clinical 12-year longitudinal study of males, the pre-capillary density was higher than post-capillary density, and the ratio was reduced from 1.39 to 1.0838. Comparing the capillary density of type 2A muscles through the vastus lateralis in subjects 19-25 and 62-72-years-of-age group confirmed a significant decrease in capillary density with age39. Another study compared the expression levels of the angiogenic inducers bFGF and PDGF in the venous wall of young and elderly patients. Both molecules were significantly reduced in the elderly40. Still another study compared the ratio of VEGF/β-actin by analyzing the gastrocnemius muscle of 22-28-year-old adults and elderly women aged 60-85 years. The ratio value was significantly reduced in the elderly women41.
Effect of Exercise on Angiogenesis During Aging
As mentioned earlier, aging is associated with reduced angiogenesis in muscle tissue and endothelial cells, with consequent reduction of angiogenic factors35. On the other hand, since exercise induces angiogenesis, it can be assumed that the exercise will lead to an improvement in angiogenesis (which could be examined in a model of aging). In relation to this, the ratio of capillary density in variable muscle type and ages was significantly increased after exercise, even when compared with the effect of exercise in younger and elderly subjects, which revealed no significant difference attributable to age42. In addition, the levels of expression of VEGF protein and RNA in human vastus lateralis biopsies were significantly increased in the elderly during exercise42. The level of VEGF was significantly elevated upon 50 minutes of mild intensity exercise in another group of elderly subjects39. In another study, eight women aged 57-76 years or 20-29 years underwent low intensity exercise four times a week for one hour. Both groups displayed similarly increased levels of vastus lateralis VEGF protein43. The collective observations indicate the age-related reduction in angiogenesis can be overcome by exercise, regardless of gender [Table 3].
Table 3.
Exercise effects in clinical studies
| Subjects | Exercise | Molecular change |
|---|---|---|
|
Young (mean age 25 years) Old (mean age 60 years) 42 |
4 days per week, with VO2 max 65 % for 8 weeks |
In vastus laterals, capillary density â VEGF RNA expressionâ |
| Old group 39 | 50 min , VO2 max 50 % of cycle ergometer | VEGFâ |
|
Aged female (57-76 years) Young female (20-29 years) 43 |
8 week, 65% Vo2 max, 1 hour 4 days each week | Old group VEGF unchanged Young group VEGF â |
Effects of Cancer and Exercise on Angiogenesis
As with exercise and aging, cancer also affects the bodyâs metabolism. In the case of cancer, angiogenesis was thought to be promoted after the formation of tumors. It is now believed that the reverse is the caseâthe tumor promoted by angiogenesis contains enlarged blood vessels, which can activate the Warburg effect1,44. Therefore, unlike the previous view, in cancer, angiogenesis plays a role in accelerating tumor progression. Despite the importance of angiogenesis, there has been little research on the relationship between cancer and exercise in terms of angiogenesis. While the effects can vary according to the type of cancer, exercise is effective in alleviating cancer45,46. A consideration of the mechanisms of cancer and exercise in terms of angiogenic factors indicates that both may induce angiogenesis, which contradicts the view that the effects of cancer are lessened by exercise47,48. These contradictory views have not been well studied. However, since exercise has a major role in maintaining homeostasis, further studies will surely be done to explore the influence of exercise on angiogenesis in cancer patients and in cancer cells.
Limitations and Further study
Angiogenesis and related phenomena have been well studied in some aspects and incompletely studied in other aspects. The latter especially includes research related to aging and cancer.
It is difficult to study the mechanism of aging due to compatibility issues with the model used. In the end, the purpose of aging model research is for clinical study. Mus musculus is a stalwart aging model. However, mice are grown in extremely limited environment, which limits the generalization of the results to aging humans49. In addition, in vivo studies are limited concerning their capacity to establish mechanisms of exercise effects on aging-induced models, so cell-based studies are needed. However, in cell-based studies, aging is defined as an accumulation of damage or errors in the cellular level and an insufficiency of self-repair50. Definitions of aging and aging model building at a cellular level are needed for studies of mechanism.
Summary
Angiogenesis has many effects on the human body. In particular, it responds to external stimuli and helps maintain homeostasis. Exercise induces angiogenesis, which in turn changes the patterns of the molecules involved. In cell-based studies, in vivo models, and clinical studies, angiogenesis is inhibited by aging and senescence, and the expressions of factors induced by angiogenesis, such as VEGF, are decreased. Although it has been shown that angiogenic factors are reduced in aging models or the elderly with exercise, there mechanism has not been elucidated in cell-based studies and in vivo models. This is because a suitable aging model has not been properly established and because the reliability of the results of studies to date can be questioned based on the characteristics of aging. In the case of cancer-related studies, most of the tissues have a tendency to display accelerated angiogenesis when tumors arise. Therefore, further studies are needed to determine how exercise affects angiogenesis in cancer. These studies are challenging because it is not easy to perform exercise with cancer patients and because of the challenges in establishing cell models or in vivo models. The establishment of proper models is necessary.




